Hûmãň Ĝéñõmĕ Ķäřŷôŧŷpè
Karyotype
A karyotype is the characteristic chromosomecomplement of a eukaryotespecies.[1][2] The preparation and study of karyotypes is part of cytogenetics.
The basic number of chromosomes in the somaticcells of an individual or a species is called the somatic number and is designated 2n. Thus, in humans 2n=46. In the germ-line(the sex cells) the chromosome number is n (humans: n=23).[1]
So, in normal diploidorganisms, autosomalchromosomes are present in two copies. There may, or may not, be sex chromosomes. Polyploidcells have multiple copies of chromosomes and haploidcells have single copies. The study of whole sets of chromosomes is sometimes known as karyology. The chromosomes are depicted (by rearranging a microphotograph) in a standard format known as a karyogram or idiogram: in pairs, ordered by size and position of centromere for chromosomes of the same size.
The study of karyotypes is made possible by staining. Usually, a suitable dye is applied after cells have been arrested during cell division by a solution of colchicine.[3] Sometimes observations may be made on non-dividing (interphase) cells. The sex of an unborn fetuscan be determined by observation of interphase cells (see amniotic centesisand Barr body).
Most (but not all) species have a standard karyotype. The normal human karyotypes contain 22 pairs of autosomal chromosomes and one pair of sex chromosomes. Normal karyotypes for womencontain two X chromosomesand are denoted 46,XX; menhave both an X and a Y chromosomedenoted 46,XY. Any variation from the standard karyotype may lead to developmental abnormalities.
Karyotypes can be used for many purposes; such as, to study chromosomal aberrations, cellularfunction, taxonomicrelationships, and to gather information about past evolutionaryevents.
Observations on karyotypes

Six different characteristics of karyotypes are usually observed and compared:[4]
- differences in absolute sizes of chromosomes. Chromosomes can vary in absolute size by as much as twenty-fold between genera of the same family: Lotus tenuis and Vicia faba (legumes), both have six pairs of chromosomes (n=6) yet V. faba chromosomes are many times larger. This feature probably reflects different amounts of DNA duplication.
- differences in the position of centromeres. This is brought about by translocations.
- differences in relative size of chromosomes can only be caused by segmental interchange of unequal lengths.
- differences in basic number of chromosomes may occur due to successive unequal translocations which finally remove all the essential genetic material from a chromosome, permitting its loss without penalty to the organism (the dislocation hypothesis). Humans have one pair fewer chromosomes than the great apes, but the genes have been mostly translocated (added) to other chromosomes.
- differences in number and position of satellites, which (when they occur) are small bodies attached to a chromosome by a thin thread.
- differences in degree and distribution of heterochromaticregions. Heterochromatin stains darker than euchromatin, indicating tighter packing, and mainly consists of genetically inactive repetitive DNA sequences.
A full account of a karyotype may therefore include the number, type, shape and banding of the chromosomes, as well as other cytogenetic information.
Variation is often found:
- between the sexes
- between the germ-lineand soma(between gametesand the rest of the body)
- between members of a population (chromosome polymorphism)
- geographical variationbetween races
- mosaicsor otherwise abnormal individuals.[5]
Error in chromosome's
Mixed up pieces
- These are called translocations, and involve pieces of non-homologous chromosomes swapping locations. They are the most common chromosomal abnormalities in humans—1 in 500 people have a translocation. See diagram to the right:
- Robertsonian translocations involve the acrocentric (chromosomes with the centromeres very near to the end) chromosomes (13, 14, 15, 21, and 22), and involves the entire chromosomes fusing together so that you are down one chromosome
- Philadelphia Chromosome: A translocation from chromosome 22 to the long arm of chromosome 9. This can lead to cancer and the abl gene is mutated
- Missing Pieces or Deletions

-
Cri-du-chat Syndrome: "Cry of the Cat" Syndrome; This is a terminal deletion: a part of the short arm near the end of chromosome 5 is deleted. The name of this deletion refers to cry of children who have this defect; they suffer from mental retardation, a shortened life span, and a distinctive facial appearance. It occurs one in 50,000 births.
- Williams Syndrome: this deletion is too small to be seen in a karyotype. You must use Fluorescence in Situ Hybridization (FISH) to find the deletion. This process works by labeling the gene in question with a fluorescent chemical (called probing). The elastin gene on chromosome 7 is deleted in this interstitial deletion (a deletion in the middle of the chromosome). This mutation brings about heart disease, since elastin is the protein that contributes to the elasticity in blood vessels.
- Other errors
- Fragile X syndrome: Look for a break, or weakness, on the long arm of the X chromosome. This causes mental retardation. It is a sex-linked factor, so more boys than girls are affected by fragile X. Since males get their X chromosome from their mother, t is not transmitted from father to son.
Learn more about another important tool used in genetic analysis: pedigrees!
Historical note
Levitsky seems to have been the first to define the karyotype as the phenotypicappearance of the somaticchromosomes, in contrast to their geniccontents.[6][7] The subsequent history of the concept can be followed in the works of Darlington[8] and White.[9][10]
Investigation into the human karyotype took many years to settle the most basic question: how many chromosomes does a normal diploidhuman cell contain?[11] In 1912, Hans von Winiwarterreported 47 chromosomes in spermatogoniaand 48 in oogonia, concluding an XX/XOsex determinationmechanism.[12] Painterin 1922 was not certain whether the diploid number of man was 46 or 48, at first favouring 46.[13] He revised his opinion later from 46 to 48, and he correctly insisted on man having an XX/XYsystem.[14] Considering their techniques, these results were quite remarkable.
New techniques were needed to definitively solve the problem:
-
- Using cells in culture
- Pretreating cells in a hypotonic solution, which swells them and spreads the chromosomes
- Arresting mitosisin metaphaseby a solution of colchicine
- Squashing the preparation on the slide forcing the chromosomes into a single plane
- Cutting up a photomicrograph and arranging the result into an indisputable karyogram.
It took until the mid 1950s until it became generally accepted that the karyotype of man included only 46 chromosomes.[15][16] Rather interestingly, the great apeshave 48 chromosomes. Human chromosome 2was formed by a merger of ancestral chromosomes, reducing the number.
Diversity and evolution of karyotypes
Although the replicationand transcriptionof DNAis highly standardized in eukaryotes, the same cannot be said for their karyotypes, which are highly variable between species in chromosome number and in detailed organization despite being constructed out of the same macromolecules. This variation provides the basis for a range of studies in what might be called evolutionary cytology.
In some cases there is even significant variation within species. In a review, Godfrey and Masters (2000) conclude: "In our view, it is unlikely that one process or the other can independently account for the wide range of karyotype structures that are observed... But used in conjunction with other phylogenetic data, karyotypic fissioning may help to explain dramatic differences in diploid numbers between closely related species, which were previously inexplicable.[17].
Changes during development
Instead of the usual gene repression, some organisms go in for large-scale elimination of heterochromatin, or other kinds of visible adjustment to the karyotype.
- Chromosome elimination. In some species, as in many sciarid flies, entire chromosomes are eliminated during development.[18]
- Chromatin diminution (founding father: Theodor Boveri). In this process, found in some copepodsand roundwormssuch as Ascaris suum, portions of the chromosomes are cast away in particular cells. This process is a carefully organised genome rearrangement where new telomeres are constructed and certain heterochromatin regions are lost.[19][20] In A. suum, all the somatic cell precursors undergo chromatin diminution.[21]
- X-inactivation. The inactivation of one X chromosome takes place during the early development of mammals (see Barr bodyand dosage compensation). In placental mammals, the inactivation is random as between the two Xs; but in marsupialsit is always the paternal X which is inactivated. Not every chromosome is normal, however some can be abnormal, resulting in a disorder when the baby is born.
Number of chromosomes in a set
A spectacular example of variability between closely related species is the muntjac, which was investigated by Kurt Benirschkeand his colleague Doris Wurster. The diploid number of the Chinese muntjac, Muntiacus reevesi, was found to be 46, all telocentric. When they looked at the karyotype of the closely related Indian muntjac, Muntiacus muntjak, they were astonished to find it had female = 6, male = 7 chromosomes.[22]
- "They simply could not believe what they saw... They kept quiet for two or three years because they thought something was wrong with their tissue culture... But when they obtained a couple more specimens they confirmed [their findings]"[23]
The number of chromosomes in the karyotype between (relatively) unrelated species is hugely variable. The low record is held by the nematodeParascaris univalens, where the haploidn = 1; the high record would be somewhere amongst the ferns, with the Adder's Tongue Fern Ophioglossum ahead with an average of 1262 chromosomes.[24] Top score for animals might be the shortnose sturgeonAcipenser brevirostrum at a mere 372 chromosomes.[25] The existence of supernumerary or B chromosomesmeans that chromosome number can vary even within one interbreeding population; and aneuploidsare another example, though in this case they would not be regarded as normal members of the population.
Ploidy: the number of sets in a karyotype
- Polyploidy, where there are more than two sets of homologous chromosomes in the cells, occurs mainly in plants. It has been of major significance in plant evolution according to Stebbins.[26][27][28][29] The proportion of flowering plants which are polyploid was estimated by Stebbins to be 30-35%, but in grasses the average is much higher, about 70%.[30] Polyploidy in lower plants (ferns, horsetailsand psilotales) is also common, and some species of ferns have reached levels of polyploidy far in excess of the highest levels known in flowering plants.
Polyploidy in animals is much less common, but it has been significant in some groups.[31]
- Haplo-diploidy, where one sex is diploid, and the other haploid. It is a common arrangement in the Hymenoptera, and in some other groups.
- Endopolyploidyoccurs when in adult differentiatedtissues the cells have ceased to divide by mitosis, but the nucleicontain more than the original somaticnumber of chromosomes.[32] In the endocycle (endomitosisor endoreduplication) chromosomes in a 'resting' nucleus undergo reduplication, the daughter chromosomes separating from each other inside an intact nuclear membrane.[33]
In many instances, endopolyploid nuclei contain tens of thousands of chromosomes (which cannot be exactly counted). The cells do not always contain exact multiples (powers of two), which is why the simple definition 'an increase in the number of chromosome sets caused by replication without cell division' is not quite accurate.
This process (especially studied in insects and some higher plants such as maize) may be a developmental strategy for increasing the productivity of tissues which are highly active in biosynthesis.[34]
The phenomenon occurs sporadically throughout the eukaryotekingdom from protozoato man; it is diverse and complex, and serves differentiationand morphogenesisin many ways.[35]
- See palaeopolyploidyfor the investigation of ancient karyotype duplications.
Species trees
The detailed study of chromosome bandingin insects with polytene chromosomescan reveal relationships between closely related species: the classic example is the study of chromosome banding in Hawaiian drosophilidsby Hampton Carson.
In about 6,500 square miles, the Hawaiian islands have the most diverse collection of drosophilid flies in the world, living from rainforests to subalpine meadows. These roughly 800 Hawaiian drosophilid species are usually assigned to two genera Drosophila and Scaptomyza in the family Drosophilidae.
The polytene banding of the 'picture wing' group, the best-studied group of Hawaiian drosophilids, enabled Carson to work out the evolutionary tree long before genome analysis was practicable. In a sense, gene arrangements are visible in the banding patterns of each chromosome. Chromosome rearrangements, especially inversions, make it possible to see which species are closely related.
The results are clear. The inversions, when plotted in tree form (and independent of all other information), show a clear "flow" of species from older to newer islands. There are also cases of colonization back to older islands, and skipping of islands, but these are much less frequent. Using K-Ardating, the present islands date from 0.4 million years ago (mya) (Mauna Kea) to 10mya (Necker). The oldest member of the Hawaiian archipelagostill above the sea is Kure Atoll, which can be dated to 30 mya. The archipelago itself (produced by the Pacific platemoving over a hot spot) has existed for far longer, at least into the Cretaceous. Previous islands now beneath the sea (guyots) form the Emperor Seamount Chain.[36]
All of the native Drosophila and Scaptomyza species in Hawaii have apparently descended from a single ancestral species that colonized the islands, probably 20 million years ago. The subsequent adaptive radiationwas spurred by a lack of competitionand a wide variety of niches. Although it would be possible for a single gravidfemale to colonise an island, it is more likely to have been a group from the same species.[37][38][39][40]
There are other animals and plants on the Hawaiian archipelago which have undergone similar, if less spectacular, adaptive radiations.[41][42]
Overview
Although much is known about karyotypes at the descriptive level, and it is clear that changes in karyotype organization has had effects on the evolutionary course of many species, it is quite unclear what the general significance might be.
- "We have a very poor understanding of the causes of karyotype evolution, despite many careful investigations... the general significance of karyotype evolution is obscure." Maynard Smith.[43]
Depiction of karyotypes
Types of banding
Cytogeneticsemploys several techniques to visualize different aspects of chromosomes:[44]
- G-bandingis obtained with Giemsastain following digestion of chromosomes with trypsin. It yields a series of lightly and darkly stained bands - the dark regions tend to be heterochromatic, late-replicating and AT rich. The light regions tend to be euchromatic, early-replicating and GC rich. This method will normally produce 300-400 bands in a normal, human genome.
- R-banding is the reverse of G-banding (the R stands for "reverse"). The dark regions are euchromatic (guanine-cytosine rich regions) and the bright regions are heterochromatic (thymine-adenine rich regions).
- C-banding: Giemsa binds to constitutive heterochromatin, so it stains centromeres.
- Q-banding is a fluorescentpattern obtained using quinacrine for staining. The pattern of bands is very similar to that seen in G-banding.
- T-banding: visualize telomeres.
Classic karyotype cytogenetics

In the "classic" (depicted) karyotype, a dye, often Giemsa(G-banding), less frequently Quinacrine, is used to stain bands on the chromosomes. Giemsa is specific for the phosphategroups of DNA. Quinacrine binds to the adenine-thymine-rich regions. Each chromosome has a characteristic banding pattern that helps to identify them; both chromosomes in a pair will have the same banding pattern.
Karyotypes are arranged with the short arm of the chromosome on top, and the long arm on the bottom. Some karyotypes call the short and long arms p and q, respectively. In addition, the differently stained regions and sub-regions are given numerical designations from proximalto distalon the chromosome arms. For example, Cri du chatsyndrome involves a deletion on the short arm of chromosome 5. It is written as 46,XX,5p-. The critical region for this syndrome is deletion of 15.2, which is written as 46,XX,del(5)(p15.2).[45]
Spectral karyotype (SKY technique)

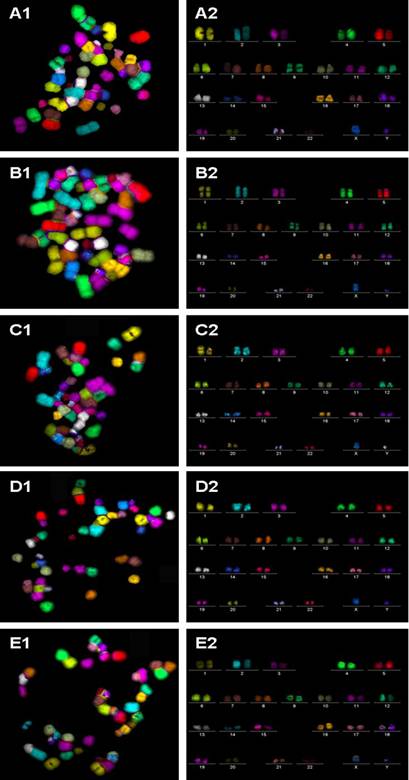
Spectral karyotyping is a molecular cytogenetictechnique used to simultaneously visualize all the pairs of chromosomesin an organism in different colors. Fluorescently-labeled probes for each chromosome are made by labeling chromosome-specific DNA with different fluorophores. Because there are a limited number of spectrally-distinct fluorophores, a combinatorial labeling method is used to generate many different colors. Spectral differences generated by combinatorial labeling are captured and analyzed by using an interferometerattached to a fluorescence microscope. Image processing software then assigns a pseudo colorto each spectrally different combination, allowing the visualization of the individually colored chromosomes.[46]
This technique is used to identify structural chromosome aberrations in cancer cells and other disease conditions when Giemsa banding or other techniques are not accurate enough.
Digital Karyotyping

{kind=link}
Digital Karyotyping is a technique used to quantify the DNA copy number on a genomic scale. Short sequences of DNA from specific loci all over the genome are isolated and enumerated. [47]
Chromosome abnormalities
Chromosome abnormalities can be numerical, as in the presence of extra or missing chromosomes, or structural, as in translocations, inversions, large-scale deletions or duplications. Numerical abnormalities, also known as aneuploidy, often occur as a result of nondisjunction during meiosis in the formation of a gamete; trisomies, in which three copies of a chromosome are present instead of the usual two, are common numerical abnormalities. Structural abnormalities often arise from errors in homologous recombination. Both types of abnormalities can occur in gametes and therefore will be present in all cells of an affected person's body, or they can occur during mitosis and give rise to a genetic mosaic individual who has some normal and some abnormal cells.
Chromosomal abnormalities that lead to disease in humans include:
- Turner syndromeresults from a single X chromosome (45, X or 45, X0).

http://www.emc.maricopa.edu/faculty/farabee/BIOBK/turner's.gif
- Klinefelter syndrome, the most common male chromosomal disease, otherwise known as 47, XXY is caused by an extra X chromosome.
- Edwards syndromeis caused by trisomy(three copies) of chromosome 18.

http://www.emc.maricopa.edu/faculty/farabee/BIOBK/kleinflt.gif
- Down syndrome, a common chromosomal disease, is caused by trisomy of chromosome 21.

http://www.emc.maricopa.edu/faculty/farabee/BIOBK/down's.gif
- Patau syndromeis caused by trisomy of chromosome 13.
- Also documented are trisomy 8, trisomy 9 and trisomy 16, although they generally do not survive to birth.
Some disorders arise from loss of just a piece of one chromosome, including
- Cri du chat(cry of the cat), from a truncated short arm on chromosome 5. The name comes from the babies' distinctive cry, caused by abnormal formation of the larynx.
- 1p36 Deletion syndrome, from the loss of part of the short arm of chromosome 1.
- Angelman syndrome? 50% of cases have a segment of the long arm of chromosome 15 missing.
Chromosomal abnormalities can also occur in cancerouscells of an otherwise genetically normal individual; one well-documented example is the Philadelphia chromosome, a translocation mutation commonly associated with chronic myelogenous leukemiaand less often with acute lymphoblastic leukemia